BiteSized Immunology: Patógenos y Enfermedades

Interacción hospedador-patógeno y evasión inmunitaria
Cualquier microorganismo que sea capaz de causar enfermedad en un organismo hospedador se denomina patógeno. Este artículo se limita a los patógenos microbianos humanos, aunque los patógenos de plantas y animales también están muy extendidos en la naturaleza. Cuando un microorganismo patógeno (bacteria, virus o parásito protozoario) infecta el cuerpo humano, se produce una batalla entre los sistemas inmunitarios innato y adaptativo del hospedador y los diversos mecanismos y factores de virulencia del patógeno. El resultado de esta batalla determina si el hospedador sobrevive y se recupera, y de qué manera. La recuperación completa implica la consecución de una homeostasis fisiológica (e inmunológica) en el hospedador, y el tiempo que esto tome dependerá de la naturaleza y la gravedad de la infección y de si ha habido alguna intervención profiláctica o terapéutica. Muchos patógenos están provistos de mecanismos de evasión inmunitaria para lograr la invasión de la célula del hospedador y su colonización, aunque también pueden utilizar con éxitos a las células de éste para acceder a los tejidos diana.
Este artículo presenta una selección de las estrategias de virulencia y de evasión inmunitaria. Entre estas últimas encontramos: (1) esconderse del sistema inmunitario (por ejemplo, dentro de las células); (2) interferir con su función (por ejemplo, señales de bloqueo) o (3) destruir algunos de sus elementos (por ejemplo, las estructuras que presentan antígenos microbianos a efectores inmunitarios para iniciar una respuesta). La virulencia generalmente implica el uso de diversos mecanismos para destruir o causar el mal funcionamiento de las células del hospedador, aunque este último también puede emplear estrategias de defensa en respuesta. Una descripción más detallada de los diversos mecanismos empleados por microorganismos específicos puede encontrarse en otras entradas de Inmunología en un mordisco.
Los virus, como Varicella zoster (varicela) y Herpesviridae (virus del herpes simple, virus Varicella-Zoster, citomegalovirus, etc.), pueden esconderse del sistema inmunitario en neuronas y células no neuronales, donde pueden persistir durante muchos años antes de aparecer en forma patogénica cuando la resistencia del hospedador disminuye. Este es el caso también de bacterias como Borrelia burgdorferi y Burkholderia pseudomallei (causantes de la enfermedad de Lyme y melioidosis, respectivamente). Hay informes de que los síntomas de la infección han reaparecido meses o años (B. burgdorferi), e incluso hasta 60 años (B. pseudomallei), después de la infección inicial. En términos de interferencia inmunitaria, existen otras estrategias más evidentes, como por ejemplo la que emplean los parásitos protozoos de Leishmania pertenecientes a Leishmania spp., que pueden inhibidor selectivamente la transcripción de citocinas proinflamatorias como interleucina 12 (IL12p40) del hospedador, lo que provoca la supresión de la respuesta inmunitaria.
Para mantener sus mecanismos de virulencia, muchas bacterias pueden secuestrar hierro libre en mamíferos, a través de la elaboración de sideróforos. El hierro es un componente esencial del metabolismo, tanto del hospedador como de los microorganismos. Para protegerse, la célula del hospedador puede sintetizar receptores de siderocalina, que compiten por la unión al hierro. Las células de mamífero también han desarrollado una serie de receptores de reconocimiento de patrones de microbios o factores microbianos, como los receptores tipo Toll (TLR), que cuando se unen a sus ligandos microbianos, desencadenan cascadas de señalización intracelular con efectos antimicrobianos.
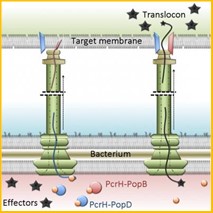
Si bien muchos patógenos bacterianos son de naturaleza intracelular, otros no necesitan invadir la célula del hospedador, sino que son capaces de secretar toxinas y otros factores de virulencia en la célula diana. Por ejemplo, hay bacterias que han desarrollado la capacidad de hacer una proyección hueca (el llamado translocón), que, al entrar en contacto con la célula del hospedador, puede transmitir factores anti-hospedador, lo que a menudo resulta en su apoptosis (llamada secreción de tipo III). Algunas de estas bacterias son Escherichia coli, Shigella flexnerii, Yersinia pestis y Chlamydia trachomatis, que causan diversos síndromes de intoxicación alimentaria, disentería, peste bubónica e infección genitourinaria, respectivamente (ver Figura 1). Sin embargo, otras bacterias, como Francisella tularensis (causante de tularemia) y Burkholderia spp. (causantes de melioidosis o muermo), tienen múltiples procesos de secreción a través de los cuales suministran factores de virulencia a la célula hospedador.
Otro patógeno humano peligroso, Bacillus anthracis, causante del ántrax, presenta mecanismos de virulencia bien desarrollados que involucran la secreción de tres proteínas, una de las cuales, el antígeno protector (AP), se une a los receptores de la célula del hospedador para facilitar la entrada del factor letal (FL) o factor edematoso (FE). Los complejos PA-LF o PA-EF entran en los endosomas tempranos a través de endocitosis mediada por clatrina, y son transportados en forma de vesículas gracias a los microtúbulos hasta los endosomas perinucleares tardíos acidificados. En estas condiciones, el FL se libera en el citoplasma, mientras que el FE permanece unido a la membrana perinuclear endosomal. En el citoplasma, el FL se escinde e inactiva a las proteína-quinasa quinasas activadas por mitógenos (MAPKK), para interrumpir la fosforilación y la transcripción en el núcleo, lo que finalmente evita la síntesis de proteínas y causa la muerte celular. Por otro lado, el FE, una adenilato ciclasa dependiente de calcio y calmodulina, causa un rápido aumento en el AMPc perinuclear, lo que resulta en edema celular, tisular y finalmente orgánico. Tanto el FL como el FE también tienen el efecto de suprimir la secreción de citocinas proinflamatorias y debilitar las barreras endoteliales vasculares al disminuir la expresión de caderina vascular, que es importante en la adhesión célula-célula; estos efectos contribuyen al aumento de la permeabilidad vascular típico del ántrax sistémico.
Los virus patógenos, por otro lado, siempre necesitan invadir una célula diana para completar sus ciclos de replicación. Uno de los más graves que han surgido en los últimos años, el virus de la inmunodeficiencia humana (VIH), afecta a las células T CD4+ y degrada la capacidad del hospedador de contraatacar con una fuerte respuesta inmunitaria mediada por células. Son tantas las tácticas de evasión inmunitaria utilizadas por el VIH, que hasta ahora han obstaculizado el desarrollo de una vacuna eficaz. En primer lugar, el VIH obtiene acceso a un individuo a través de sitios inmunológicamente privilegiados, como la vagina o el recto, que no están bien provistos de tejido linfoide; posteriormente el virus utiliza correceptores (CCR5/ CXCr4) para obtener acceso a las células T CD4+. El resultado es una pérdida gradual de éstas últimas células, lo que provoca el deterioro de la respuesta inmunitaria mediada por células y un incremento de la susceptibilidad a otras infecciones (neumonía bacteriana) o tumores (por ej. Sarcoma de Kaposi). Los casos de VIH pueden controlarse con medicamentos antirretrovirales mientras se identifican y evalúan algunas vacunas. Variola major (causante de la viruela) también ha desarrollado tácticas de evasión inmunitaria, al secretar una proteína que inhibe la activación de las enzimas del complemento y otra, la proteína de unión a quimiocinas de tipo II, que bloquea las señales de reclutamiento de células del sistema inmunitario e inflamación en el sitio de la infección. Los patógenos también pueden evadir el sistema de procesamiento antigénico y presentación del hospedador, por ejemplo, al interferir con la expresión de las proteínas de membrana de MHC de clase I, aunque esto a su vez aumenta su susceptibilidad a la actividad de las células asesinas naturales (NK, por sus siglas en inglés). Sin embargo, los miembros de las familias de herpesvirus, papilomavirus, retrovirus, poxvirus y flavivirus también han desarrollado mecanismos para escapar del ataque de las células NK y promover su propia supervivencia en las células del hospedador mediante la inhibición de la apoptosis en estas células. Finalmente, algunos patógenos cambian con frecuencia los antígenos de superficie que muestran, siendo el virus de la influenza (ortomixovirus) un excelente ejemplo de esto.
A través de todos estos mecanismos de virulencia y técnicas de evasión inmunitaria, los patógenos han desarrollado estrategias para sobrevivir en el hospedador. Por supuesto, la interacción de un patógeno con un hospedador es una situación dinámica, por lo que los avances realizados por el patógeno son contrarrestados por el hospedador. Por lo tanto, la expansión de las células T citotóxicas (Tc) en el hospedador puede inducir a mutaciones “de escape” en el patógeno para evitar ser un objetivo futuro de las Tc. Por lo tanto, los patógenos continuarán evolucionando y emergiendo, y posiblemente los más exitosos son aquellos que explotan a sus anfitriones sin matarlos.
La versión en español ha sido coordinada por Jesús Gil-Pulido, responsable de redes sociales de la Sociedad Española de Inmunología, y ha sido posible gracias a miembros de la Sociedad Española de Inmunología (https://www.inmunologia.org/index.php)
© The copyright for this work resides with the author.