BiteSized Immunology: Patógenos y Enfermedades

Virus de la inmunodeficiencia humana (VIH)
El virus de la inmunodeficiencia humana (VIH), un miembro de la familia de los retrovirus, es el agente causante del síndrome de inmunodeficiencia adquirida (SIDA). El VIH invade varias células del sistema inmunitario (por ejemplo, células T CD4+ y monocitos), lo que resulta en una disminución del número de células T CD4+ por debajo del nivel crítico y la pérdida de inmunidad mediada por células, de modo que el organismo se vuelve cada vez más susceptible a las infecciones oportunistas y al cáncer.
Invasión de las células del sistema inmunitario por el VIH
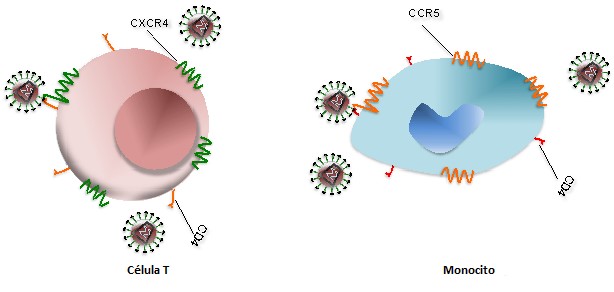
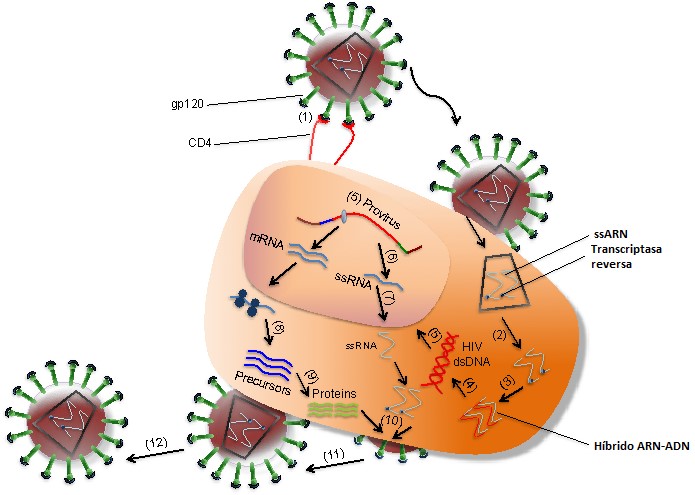
El VIH infecta a las células T a través de la interacción de alta afinidad entre la glicoproteína de la envoltura del virión (gp120) y la molécula CD4. Durante la infección también interviene el correceptor de células T, llamado CXCR4, mientras que, para la infección de los monocitos, el VIH interactúa con el correceptor CCR5 (Figura 1). Como se ilustra en la Figura 2, después de que gp120 se una a CD4 en la célula T (1), las nucleocápsidas que contienen el genoma viral y las enzimas entran en la célula diana (2). Tras la liberación del genoma viral y las enzimas de la cápsida, la transcriptasa reversa viral cataliza una reacción a partir del ARN de cadena simple (ssARN) para formar híbridos de ARN-ADN (3). Para producir ADN de doble cadena (dsADN), la cadena de ARN viral es degradada parcialmente por la ribonucleasa H, lo que permite la síntesis de la segunda cadena de ADN (4). El dsADN viral se traslada al núcleo y se integra en el genoma del huésped por la acción de la integrasa viral (5). Los factores de transcripción transcriben el ADN proviral en ssARN genómico (6), que se exporta al citoplasma (7). Una vez ahí, los ribosomas de la célula infectada catalizan la síntesis de proteínas virales precursoras (8), que se escinden en proteínas virales mediante las proteasas del virus (9). El ssARN y las proteínas del VIH se ensamblan debajo de la membrana plasmática de la célula infectada (10), formando nuevos viriones a partir de ella (11). La maduración se produce ya sea en las yemas de formación o después de la brotación de la célula infectada (12). Durante la maduración, las proteasas del VIH rompen las poliproteínas en proteínas de VIH funcionales individuales. Los viriones maduros son capaces de infectar otra célula.
Respuesta inmunitaria innata frente al VIH
Las células del sistema inmunitario innato (por ejemplo, las células dendríticas y las células asesinas naturales) son la primera línea de defensa que encuentra el VIH al ingresar al organismo.
Macrófagos. Los macrófagos tisulares son una de las células diana del VIH. Estos macrófagos son portadores del virus y se sabe que son una fuente de proteínas virales. Sin embargo, se ha demostrado que los macrófagos infectados pierden su capacidad para ingerir y matar microbios extraños y presentar antígeno a las células T. Esto podría suponer una gran parte de la disfunción inmunitaria general causada por la infección por VIH.
Células dendríticas (DCs, por sus siglas en inglés). Las DCs son células grandes con extensiones citoplásmicas tipo dendritas. Estas células presentan antígenos procesados a los linfocitos T en los ganglios linfáticos. Las DCs epidérmicas, que expresan CD1a y gránulos de Birbeck, probablemente se encuentran entre las primeras células inmunitarias que combaten el VIH en las superficies mucosas. Estas células transportan el VIH desde el sitio de la infección hasta el tejido linfoide. Las DCs foliculares, que se encuentran en el tejido linfoide, son también células presentadoras de antígeno, capaces de atrapar antígenos y presentarlos en su superficie celular. En los folículos de los ganglios linfáticos, las DCs proporcionan señales para la activación de los linfocitos B.
Células asesinas naturales (NK, por sus siglas en inglés). Las células NK tienen actividad lítica contra células con expresión disminuida de antígenos de clase I del complejo principal de histocompatibilidad (MHC, por sus siglas en inglés). Debido a que se requiere la presencia de MHC de clase I para la presentación de péptidos a los receptores de células T, las células NK son una importante línea de defensa cuando el VIH se escapa de la respuesta inmunitaria celular. Las células NK proliferan en respuesta al interferón tipo 1 secretado por las DCs. Estas células NK estimuladas liberan citocinas, como el interferón γ (IFN-γ), el factor de necrosis tumoral α (TNF-α) y las quimiocinas para activar la proliferación de células T (respuesta inmune celular). Las células NK también inhiben la replicación viral al liberar IFN-γ.
Respuesta inmunitaria adaptativa frente al VIH
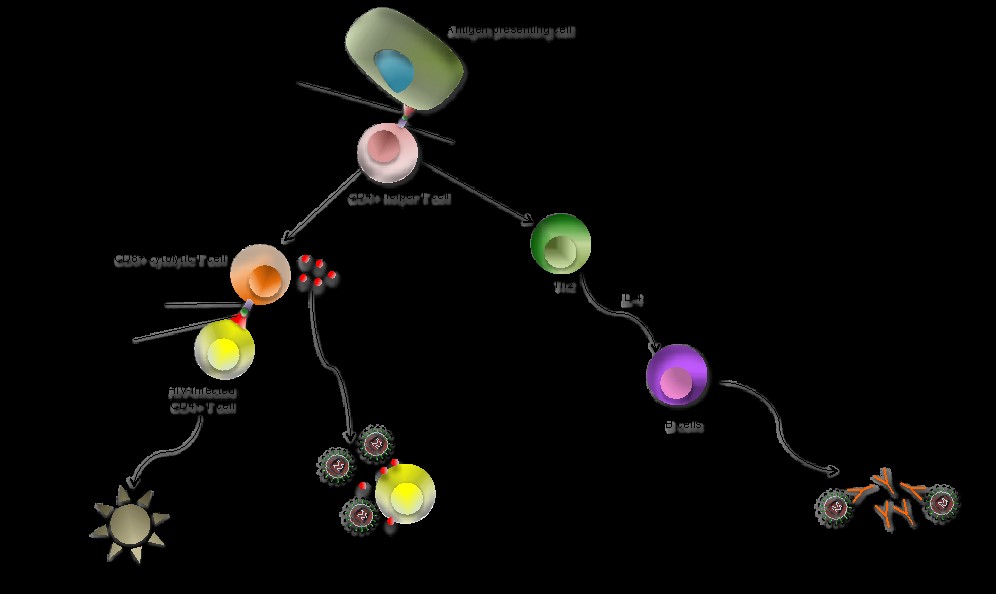
Respuesta inmune celular. La respuesta inmune celular es inducida tras la entrada del VIH en las células diana (por ejemplo, células T) y la síntesis de proteínas virales (Figura 1). Las moléculas de MHC de clase I de la superficie celular presentan los fragmentos peptídicos del VIH degradados intracelularmente para su reconocimiento por los receptores de células T (TCR, por sus siglas en inglés) de los linfocitos T CD8+ (Figura 3). Las células T CD8+ lisan las células infectadas por el VIH y secretan citocinas, cómo interferón-γ (IFN-γ), factor de necrosis tumoral α (TNF-α) y quimiocinas tales como MIP-1 α, MIP β y RANTES, que inhiben la replicación del virus y bloquean la entrada viral en células T CD4+. El desarrollo de células T CD8+ es crucial para el control de la replicación del VIH. Esto resulta en una disminución de la viremia después de la infección primaria. En las primeras etapas de la infección, las células T CD4+ pierden su capacidad proliferativa y, por lo tanto, su contribución al control viral es menor. Sin embargo, durante la infección crónica, las células T CD4 + están presentes y secretan interleucina-2 (IL-2) o citocinas, como el IFN-γ, para controlar la viremia.
Respuesta humoral. La respuesta inmunitaria humoral ocurre más tarde, por lo tanto, el nivel de anticuerpos durante la infección aguda es muy bajo. Los anticuerpos no neutralizantes para proteínas estructurales (P17 y P24) aparecen primero y generalmente no persisten. Posteriormente, se generan anticuerpos neutralizantes específicos para las proteínas implicadas en la entrada del virus a las células. Estos anticuerpos son específicos para: (1) la región variable de gp120 (V3); (2) sitios de unión a CD4 y receptores de quimiocinas (es decir, CXCR4 y CCR5); (3) la proteína transmembrana gp41. Se ha observado que algunos anticuerpos neutralizantes son muy potentes y juegan un papel clave en el control de la infección por VIH en unos pocos individuos VIH+ sin síntomas que mantienen alto el nivel de células T CD4+ y una carga viral baja.
¿Por qué el sistema inmunitario no combate al VIH?
Hay varias razones que pueden contribuir a que el sistema inmunitario no controle la infección por VIH y prevenga el desarrollo del SIDA. Al infectar las células T CD4+, el VIH puede replicarse en las células T activadas y paralizar uno de los componentes principales del sistema inmunitario adaptativo. El VIH también puede establecer una infección latente en las células T CD4+ y permanecer invisible para las células T CD8+, por lo que la replicación del virus para generar nuevos viriones puede ocurrir con posterioridad a la infección. La mutación antigénica que afecta a los epítopos reconocidos por las células T, puede afectar a la capacidad de unión de las moléculas MHC a los péptidos virales, lo que resulta en la incapacidad de los TCR para reconocer el complejo péptido-MHC. Finalmente, el VIH es capaz de ocultarse de los anticuerpos anti-VIH mediante la expresión de glicanos no inmunogénicos sobre epítopos antigénicos clave.
La versión en español ha sido coordinada por Jesús Gil-Pulido, responsable de redes sociales de la Sociedad Española de Inmunología, y ha sido posible gracias a miembros de la Sociedad Española de Inmunología (https://www.inmunologia.org/index.php)
© The copyright for this work resides with the BSI
.