BiteSized Immunology: Receptores y Moléculas

Las uniones Gap en el sistema inmunitario
Introducción
Las uniones intercelulares tipo Gap (GJs) son el mecanismo mejor estudiado para el establecimiento de contactos estrechos entre células. Aunque las células del sistema inmunitario normalmente están diseminadas por los tejidos, pueden “tocarse” unas a otras durante las respuestas inmunitarias o en situaciones patológicas, como en leucemias, en donde los órganos linfoides están infiltrados por células que proliferan y que son clonalmente idénticas. En este caso las células pueden contactar a través de las GJs.
¿Qué son las uniones Gap?
Las GJ están formadas por dos conexones acoplados (hemicanales), cada una perteneciente a una célula vecina. Cada conexón individual está compuesta por seis subunidades oligomerizadas de la proteína conexina (Cx), perteneciente a una familia de proteínas transmembrana relacionadas estructuralmente. Todas las Cxs muestran una estructura similar, que se caracteriza por cuatro dominios hidrofóbicos formados por hélices alfa transmembrana (ver figura). A día de hoy, existen miembros de la familia de Cx altamente conservados tanto en el genoma de humanos (alrededor de 20) como en el de ratón (alrededor de 21), algunos de los cuáles se expresan en células del sistema inmunitario (Tabla 1). La formación de las GJs permite la transferencia de forma directa de iones, de moléculas pequeñas y también de miRNAs entre células adyacentes. Por ejemplo, las células del miocardio están conectadas por GJs que permiten su polarización de manera simultánea. Se conocen tres tipos diferentes de GJs: homoméricas/homotípicas, heteroméricas y heterotípicas. Las uniones homotípicas están formadas por dos conexones idénticos, mientras que las heterotípicas están formadas por diferentes conexones. Los hemicanales homoméricos y heteroméricos contienen uno o más isotipos de conexinas.
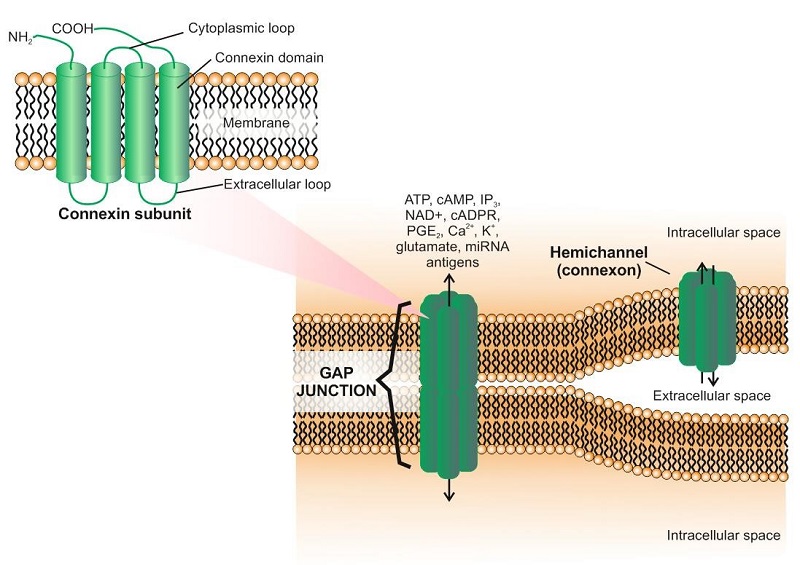
El papel de las conexinas está limitado en algunas ocasiones a la señalización intracelular, algo que no requiere necesariamente la formación de GJs funcionales (Figura). En este caso, se activan mecanismos de señalización intracelular en los que están implicadas moléculas como ZO-1 y otras. Este mecanismo puede funcionar tanto en situaciones fisiológicas como patológicas.
Recientemente se ha descrito otra familia de proteínas, llamadas panexinas (Panxs) y formada por solo tres miembros (Panx1-3), con la capacidad de formar GJs. Sin embargo, la funcionalidad de GJs formados por Panx es controvertida.

¿Cuál es la función de las GJs en el sistema inmunitario?
Muchas moléculas inmunológicamente relevantes pasan a través de las uniones GJs, incluyendo péptidos pequeños como IP3, cAMP o el Ca2+. Este último juega un papel importante en las células del sistema inmunitario, ya que contribuye a la migración, maduración y muerte celular. Hace ya muchos años que se demostró la transferencia de segundos mensajeros asociados a la señalización por Ca2+, como es el IP3, a través de las uniones GJs.
Las uniones GJs en inmunología atrajeron la atención de los investigadores cuando se demostró la transferencia de antígenos a través de estas en células presentadoras de antígeno. Este tipo de transferencia permite la presentación cruzada de antígenos en complejos con moléculas de MHC de clase I. En este proceso, las células presentadoras de antígeno adquieren antígenos provenientes de células infectadas o tumorales e inician una respuesta inmunitaria presentándolos a las células T. De hecho, las conexinas se expresan en todos los tipos de células presentadoras de antígenos. También se ha demostrado la presencia de uniones GJ entre células T polarizadas (Th1 o Th2)
Las células dendríticas foliculares (FDCs) son células especializadas que retienen antígenos y forman parte tanto del establecimiento como del mantenimiento de la respuesta inmunitaria. Se ha demostrado que Cx23 colocaliza con los marcadores FDC (CD21 y CD35) en los centros germinales de las amígdalas y del bazo en humanos. Además, también se ha demostrado comunicación por uniones GJ entre FDCs y entre FDCs y células B a nivel funcional y ultraestructural.
El NAD+ es una pequeña molécula capaz de permear las uniones GJs y los hemicanales; por tanto, no es de extrañar que la transferencia o liberación de NAD+ a través de estos canales contribuya a la comunicación entre diferentes células del sistema inmunitario. Por ejemplo, la molécula CD38 se expresa en células mieloides y linfoides que usan el NAD+ para generar cADPR (por sus siglas en inglés) y ribosa de ADP, los cuales participan en multitud de respuestas inmunitarias.
El ATP activa a las células del sistema inmunitario y también participar en respuestas autocrinas y paracrinas cuando es liberado por las células. Se ha demostrado recientemente la contribución de los hemicanales de Cx y Panx a la señalización purinérgica. Por lo tanto, los hemicanales de Cx y Panx median la liberación de ATP, que podría tener un papel en todos los pasos de la respuesta inmunitaria. El cAMP también puede ser transferido a través de las uniones GJs, contribuyendo a la inmunosupresión mediada por las células T reguladoras cuando entran en contacto con las células T naive.
En ensayos in vitro se ha observado que las células T CD8+ usan las uniones GJs para su activación. Por ejemplo, las células T CD8+ naive establecen uniones GJs con las células de melanoma. Aunque las células T CD8+ no usan las GJs para destruir, las células NK sí que establecen uniones GJs con las células dendríticas y con las céluls tumorales en un proceso Cx43-dependiente que contribuye a la lisis de éstas últimas y, por lo tanto, a la inmunidad antitumoral.
Las uniones GJs permiten el transporte entre células de moléculas de ARN de cadena simple o de doble cadena, así como de microRNAs de cadena simple, teniendo todo ello un alto impacto sobre la respuesta inmunitaria. Recientemente se ha demostrado la transferencia de diferentes microRNAs (miR-142, -223, -155) entre el mismo tipo de células del sistema inmunitario. De la misma manera se ha demostrado que los macrófagos inhiben la proliferación de las células del carcinoma hepático mediante la transferencia de miRNA a través de las uniones GJs. Es muy interesante el hecho de que solo algunos miRNAs específicos son los que se intercambian entre las células, lo que indica la existencia de una permeabilidad selectiva de las uniones GJs para ciertos miRNAs. Estos datos sugieren que la transferencia de miRNAs podría ser un mecanismo del sistema inmunitario para combatir el cáncer.
En caso de enfermedad, las uniones GJs parecen tener un importante papel en algunos casos, como las leucemias. En algunos desórdenes hematológicos se ha observado un aumento de la expresión de Cx43 en células del estroma de la médula ósea, como es el caso de la leucemia linfoblástica aguda, la mieloide (ALL y AML, por sus siglas en inglés) y en algunos desórdenes crónicos mieloproliferativos. Sin embargo, en leucemia aguda también existen datos que indican una reducción de la expresión de Cx43 y una pérdida de comunicación por GJ entre las células del estroma y las células leucémias. También se ha observado en algunos tipos de leucemias que las uniones GJs son cruciales para la supervivencia celular, por ejemplo, activando señales de supervivencia de las células madre de la leucemia (LSCs, por sus siglas en inglés). Por todo ello, parece claro que la actividad de las células leucémicas es altamente dependiente de la comunicación con otras células del estroma a través de las uniones GJs. A pesar de todo, todavía se desconoce el significado de las variaciones en la expresión de Cx43 en la patogénesis de estas enfermedades.
¿Cómo se regulan las uniones Gap?
Las uniones GJs se pueden formar, abrir, cerrar y desaparecer muy rápido. Esta plasticidad les permite mediar comunicaciones celulares tanto lentas como rápidas, pero también indica que este proceso debe estar regulado de forma precisa. Los hemicanales, como norma, se forman completamente en la membrana del retículo endoplásmico y se transportan a través del complejo de Golgi a la superficie celular. Las Cxs están siendo continuamente formadas y degradadas en el lisosoma después de su internalización por endocitosis. La degradación de las placas de las uniones GJs se regula mediante ubiquitinización y fosforilación y, además en algunos tipos celulares, es un mecanismo dependiente del proteosoma.
La expresión de las conexinas está regulada por numerosas citocinas, las cuales normalmente no actúan solas, sino que tienen acciones sinérgicas (ver tabla 2).

¿Cuáles son las implicaciones clínicas de los estudios de las uniones GJ?
Existe la posibilidad de usar drogas específicas para inhibir las uniones GJs y los hemicanales. Esto puede llevarse a cabo con el uso de bloqueantes no específicos de las uniones GJs, como son el octanol o la carbenoxolona, o con el uso de moléculas más específicas, como son los péptidos miméticos. En la actualidad están surgiendo nuevas aproximaciones, como son el uso de anticuerpos frente a las conexinas. El descubrimiento de la posible transferencia de miRNAs a través de las uniones GJs abre nuevas posibilidades de actuación sobre microRNAs y siRNA específicos. Todo ello puede abrir nuevas oportunidades para ajustar la respuesta inmunitaria innata y adaptativa y tratar algunas enfermedades, como la leucemia.
References
1. Regulation of Hemichannels and Gap Junction Channels by Cytokines in Antigen-Presenting cells. Saez PJ, Shoji KF, Aguirre A, et al. Mediators Inflamm. 2014;2014:742734.
2. Cell communication across gap junctions: a historical perspective and current developments. Evans WH. Biochem Soc Trans. 2015;43(3):450-9.
3. Cx25 contributes to leukemia cell communication and chemosensitivity. Sinyuk M, Alvarado AG, Nemiyanov P, et al. Oncotarget. 2015;6(31):31508-21.
4. ZO-1 expression shows prognostic value in chronic B cell leukemia. Nesmiyanov PP, Tolkachev BE, Strygin AV. Immunobiology. 2016;221(1):6-11.
5. Gap junctional shuttling of miRNA – A novel pathway of intercellular gene regulation and its prospects in clinical application. Lemcke H., Steinhof G., David R. Cellular Signalling.2015;27:2506-2514.
© The copyright for this work resides with the BSI