BiteSized Immunology: Sistemas y Procesos

Sistema del complemento
El complemento fue descubierto por Jules Bordet como un componente termolábil del plasma normal que causaba la opsonización y destrucción de las bacterias. El sistema del complemento hace referencia a un conjunto de más de 20 proteínas que circulan por la sangre y los fluidos tisulares. Muchas de estas proteínas están normalmente inactivas, pero en respuesta al reconocimiento de componentes moleculares en los microorganismos, pueden activarse secuencialmente a través de una cascada enzimática (la activación de una proteína favorece la fragmentación y activación de la siguiente proteína de la ruta). El complemento puede ser activado a través de tres rutas distintas (Figura 1), causando la activación de C3, su fragmentación en un fragmento grande, C3b que actúa como opsonina, y otro pequeño, C3a (una anafilotoxina) que promueve inflamación. C3 activado puede promover la vía lítica, que es capaz de dañar las membranas plasmáticas de las células y algunas bacterias. C5a, producida en este proceso, atrae macrófagos, neutrófilos y algunos mastocitos.
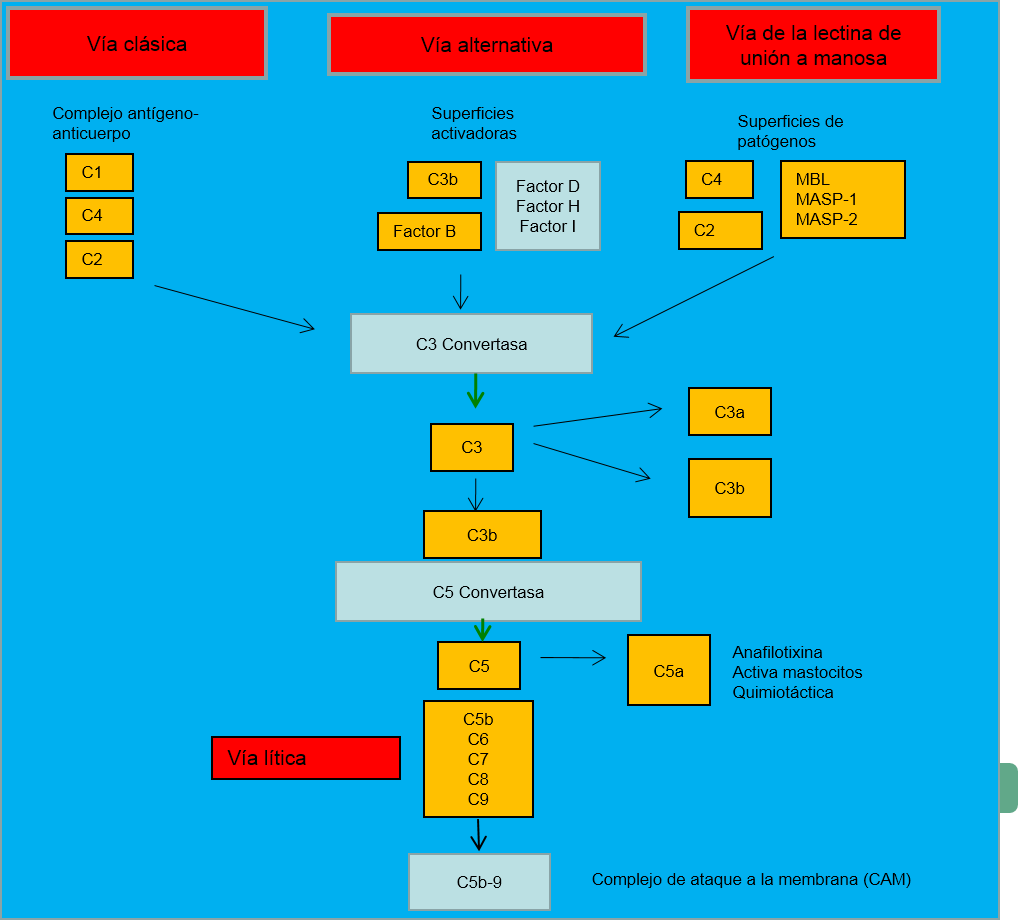
Vía clásica
Esta ruta involucra los componentes C1, C2 y C4. Se inicia por complejos antígeno-anticuerpo unidos a C1, que de por sí cuenta con tres subcomponentes, C1q, C1r y C1s. La vía activa la convertasa C3, C4b2a, que permite la fragmentación de C3 en dos fragmentos: el grande (C3b), que puede unirse covalentemente a la superficie de patógenos microbianos y opsonizarlos, y el pequeño (C3a), que activa mastocitos, lo que causa la liberación de mediadores vasoactivos como la histamina.
Vía alternativa
Esta ruta involucra a los factores B, D, H e I, que interactúan entre sí, y C3b, permitiendo la generación de la convertasa C3, C3bBb, que puede activar más C3; esta es la razón por la que muchas veces se la conoce como “bucle de amplificación”. La activación del bucle tiene lugar en presencia de paredes bacterianas y fúngicas, pero se inhibe por ciertas moléculas presentes en la superficie de células normales de mamíferos.
Vía de la lectina de unión a manosa
Esta ruta es activada por la unión de lectinas de unión a manosa (MBL) a residuos de manosa en las superficies de patógenos. Como consencuencia se activan las serín proteasas asociadas a MBL, MASP-1 y MASP-2, que activan C4 y C2 para formar la convertasa C3, C4b2a.
Vía lítica
Esta ruta se inicia tras la escisión de C5, y la unión de C5b a su diana. C6, C7, C8 y C9 se unen a C5b formando el complejo de ataque a la membrana (CAM), que, cuando se inserta en la membrana externa de algunas bacterias, facilita su muerte por lisis. Las células sanguíneas que tienen anticuerpos unidos a su superficie pueden también activar la vía clásica y lítica y ser susceptibles a la lisis por este mecanismo.
Papel del complemento en la enfermedad
El sistema del complemento juega un papel crítico en la inflamación y defensa contra algunas infecciones bacterianas. Además, puede activarse tras trasfusiones sanguíneas incompatibles, y durante las respuestas inmunitarias dañinas que acompañan a las enfermedades autoinmunitarias. Las deficiencias individuales de los componentes del complemento o los inhibidores del sistema, pueden dar lugar a una gran variedad de patologías (Tabla 1), lo que indica su importancia en la protección frente a la enfermedad.
Tabla 1. Enfermedades asociadas con deficiencias en el complemento
| Deficiencia | Enfermedad |
| C3 y factor B | Infecciones bacterianas graves |
| C3b-INA, C6 y C8 | Infecciones por Neissaria graves |
| Deficiencia de los componentes C tempranos, C1, C4 y C2. | Lupus sistémico eritomatoso (SLE), glomerulonefritis y polimiositis. |
| Inhibidor de C1 | Angioedema hereditario |
La versión en español ha sido coordinada por Jesús Gil-Pulido, responsable de redes sociales de la Sociedad Española de Inmunología, y ha sido posible gracias a miembros de la Sociedad Española de Inmunología (https://www.inmunologia.org/index.php).
© The copyright for this work resides with the author