BiteSized Immunology: Receptors & Molecules

Gap junctions in the immune system
Introduction
Gap Junctions (GJs) are the most studied mechanism of cell-cell close contact communications. Though immune cells are generally sparse within tissues, they can ‘touch’ each other during normal immune response or in pathological conditions, such as leukemia, when lymphoid organs are tightly infiltrated by clonally identical proliferating cells. In this case they can contact through GJs.
What are Gap Junctions?
GJ channels consist of two docked connexons (hemichannels), each provided by one of the neighbouring cell. A single hemichannel is composed of six oligomerized Connexin (Cx) subunits which belong to a family of structurally related transmembrane proteins. All Cxs share a similar structure that is characterized by four alpha helical transmembrane hydrophobic domains (see Figure). Today, there are 20 and 21 highly conserved members of the Cx family in the human and mouse genomes respectively, some of which are expressed on the immune cells (see Table 1). Formation of GJs allows direct transfer of ions, small molecules and miRNAs between the adjacent cells. Myocardial cells are connected by GJs which allows their simultaneous polarization. Three different types of GJs are known: homomeric/homotypic, heteromeric and heterotypic. Homotypic GJs are formed by two identical connexons, while heterotypic are formed by different connexons. Homomeric or heteromeric hemichannels comprise one or more Cx isotypes.
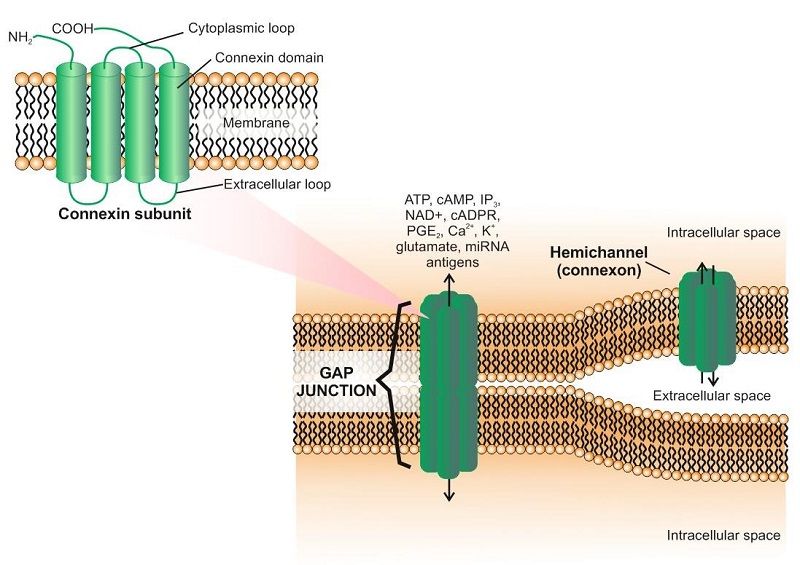
The role of connexins is sometimes limited to intracellular signaling, which does not necessary require formation of functional GJs (Figure). In this case, intracellular signaling pathways are activated, involving such molecules as ZO-1 and others. This mechanism may be employed both in normal and pathological states.
Recently, another family of proteins named Pannexins (Panxs) comprising of only three members (Panx1–3) was proposed to form GJs. However, the expression of functional Panx GJs still remains controversial.
What are the functions of GJs in the immune system?
Many immunorelevant molecules can permeate GJs. They include small peptides, IP3, cAMP. Ca2+ signalling plays a major role in immune cells, contributing to cell migration, maturation, and death. The transference of second messengers associated with Ca2+ signalling, such as IP3, through GJs was demonstrated several years ago.
GJs in immunology have gained attention when the antigen transfer through GJs in APCs was demonstrated. This direct antigen transfer through GJs allows antigen cross-presentation in complex with MHC class I molecules. In this process antigen-presenting cells acquire antigens from infected or tumor cells and initiate an effective immune response by presenting antigens to T cells. Connexins are in fact expressed in all types of antigen-presenting cells. GJs between polarized T cells (Th1 orTh2) have also been demonstrated.
Follicular dendritic cells are specialized cells that retain antigens and take part in the immune response formation and sustainment. It was demonstrated that Cx43 colocalizes with FDC markers (CD21 and CD35) at germinal centers of human tonsils and spleen. Moreover, GJ communication among FDCs and between FDCs and B cells has been demonstrated at functional and ultrastructural levels.
NAD+ is a small molecule that permeates GJs and hemichannels; it can be anticipated that NAD+ transfer or release through these channels might contribute to cell-cell communication in different immune cells. For example, CD38 is a molecule expressed by myeloid and lymphoid cells that use NAD+ to generate cADPR and ADPribose, which contribute to several immune cell responses.
ATP activates immune cells and also contributes to autocrine and paracrine activation when released from cells. The contribution of Cx and Panx hemichannels to purinergic signalling has been reported and was recently revised. Thus, Cx and Panx hemichannel-mediated ATP release might play a role in all steps of the immune response. cAMP also can be transferred through GJs; this contributes to immunosuppression mediated by Tregs when they come into contact with naïve T cells.
CD8+ cells use GJs for activation which was shown in vitro: naive CD8+ T cells establish GJs with melanoma target cells. CD8+ cells do not use GJs for killing; however, NK cells establish GJs with DCs and tumor cells in a Cx43-dependent process that contributes to NK cell-mediated lysis and further antitumor immunity.
GJs allow cell-cell shuttling of single- and double-stranded RNA, as well as specific single-stranded microRNA, which has a high impact on immune responses. Recently, the cell-cell transfer of different microRNAs (miR-142, -223, -155) between identical immune cells was demonstrated. Likewise, macrophages were found to inhibit proliferation of hepatic carcinoma cells by transferring miRNA through GJs. Interestingly, only specific miRNAs are exchanged between cells, indicating a selective permeability of GJs for certain miRNAs. These data suggest miRNA transfer as a mechanism of cancer repression by the immune system.
In disease, GJs have been shown to play a certain role in leukemia. Upregulation of Cx43 was found in stromal bone marrow cells in hematologic disorders, including acute lymphoblastic and myeloid leukemia (ALL and AML) and chronic myeloproliferative disorders. However, in acute leukemia, there is also evidence of a reduction in Cx43 expression and loss of GJ communication between stromal and leukemic cells. It was also shown that GJs are crucial for cell survival in some leukemia types, specifically providing survival signal for leukemia stem cells (LSCs). Thus, it seems clear that the activity of leukemic cells is strongly dependent on communication with each other and with stromal cells through GJs, however, the significance of variation in Cx43 expression in the pathogenesis of these diseases is yet to be explored.
How Gap Junctions are regulated?
GJs can be quickly formed, open/close and be quickly disrupted. This plasticity allows them to mediate both slow and fast intercellular communications but at the same time that means that GJ functions should be precisely regulated. Hemichannels, as a rule, are fully formed in the membranes of the endoplasmic reticulum and transported through the Golgi complex to the cellular surface. Cxs are continuously renewed and degraded by the lysosome after internalization through endocytosis. Degradation of GJ plaques is regulated by ubiquitination and phosphorylation and is also proteasome dependent in some cell types.
Expression of Cxs is regulated by several cytokines which don’t usually act alone but have a synergic actions (see table 2).

What are the clinical implications of GJ studies?
There is a possibility for the use of specific drug delivery to inhibit GJs and hemichannels. This can be achieved by the use of non-specific GJ blockers, such as octanol or carbenoxolone or by using more specific molecules, such as mimetic peptides. New approaches are rising, such as using antibodies against connexins. The data on a miRNA transfer through GJs open new possibilities for the specific targeting of microRNA and siRNA. Altogether, it can open new possibilities to adjust the innate and adaptive immune responses and to treat some diseases, such as leukemia.
References
1. Regulation of Hemichannels and Gap Junction Channels by Cytokines in Antigen-Presenting cells. Saez PJ, Shoji KF, Aguirre A, et al. Mediators Inflamm. 2014;2014:742734.
2. Cell communication across gap junctions: a historical perspective and current developments. Evans WH. Biochem Soc Trans. 2015;43(3):450-9.
3. Cx25 contributes to leukemia cell communication and chemosensitivity. Sinyuk M, Alvarado AG, Nemiyanov P, et al. Oncotarget. 2015;6(31):31508-21.
4. ZO-1 expression shows prognostic value in chronic B cell leukemia. Nesmiyanov PP, Tolkachev BE, Strygin AV. Immunobiology. 2016;221(1):6-11.
5. Gap junctional shuttling of miRNA – A novel pathway of intercellular gene regulation and its prospects in clinical application. Lemcke H., Steinhof G., David R. Cellular Signalling.2015;27:2506-2514.
© The copyright for this work resides with the BSI