BiteSized Immunology: Experimental Techniques

Production of MHC Class I Tetramers
Major histocompatibility complex (MHC) tetramers were first described in the mid-1990s, and since then have been an invaluable tool to all T-cell immunologists. Identifying antigen-specific T cells is inherently difficult due to the low avidityof the interaction between T cell receptor (TCR) and MHC:peptide complex. The interaction has a half life of a mere 10 seconds!
The production of MHC class I tetramers is shown in Figure 1. First MHC heavy chains expressing a bacterial BirA-recognition site are synthesised. The MHC heavy chain is then folded with β2-microglobulin (β2M; the MHC light chain) and synthetic peptide. The enzyme BirA is then added to biotinylate the complex, adding a biotin molecule to each MHC monomer that is formed. In the presence of streptavidin which has four biotin binding sites per molecule, four MHC monomers are joined together to form a tetramer.
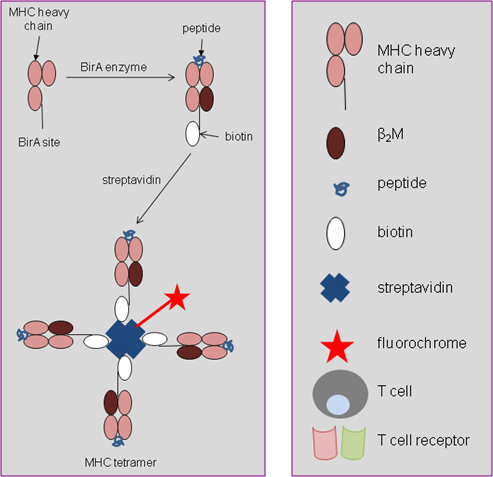
Whilst single MHC:peptide complexes and TCR bind weakly, MHC tetramers can bind up to four TCRs simultaneously, creating a much stronger interaction. MHC tetramers only bind to the relevant TCR (Figure 2), and tetramers specific for any MHC heavy chain (e.g. HLA A2, HLA B7) can be produced, coupled with any peptide epitope of choice (e.g. viral, bacterial, tumour and autoimmune antigens).
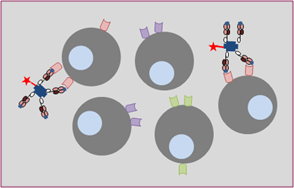
The streptavidin molecule is routinely linked to a fluorochrome such as phycoerythrin, to allow
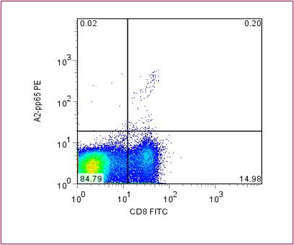
for direct staining for use with flow cytometry. An example of tetramer staining is shown in Figure 3; here peripheral blood mononuclear cells (PBMCs) have been stained with an antibody specific for CD8 and also a tetramer of HLA-A2 molecules containing a cytomegalovirus (CMV) peptide.
It can be seen that 1.3% of CD8+ cells are tetramer positive. Antigen-specific T cells as low as 0.05% of all T cells can be measured.
© The copyright for this work resides with the author