BiteSized Immunology: Pathogens & Disease

Human Immunodeficiency Virus (HIV)
Human immunodeficiency virus (HIV), a member of the retrovirus family, is the causative agent of acquired immunodeficiency syndrome (AIDS). HIV invades various immune cells (e.g., CD4+ T cells and monocytes) resulting in a decline in CD4+ T cell numbers below the critical level, and loss of cell-mediated immunity − therefore, the body becomes progressively more susceptible to opportunistic infections and cancer.
HIV invasion of immune cells
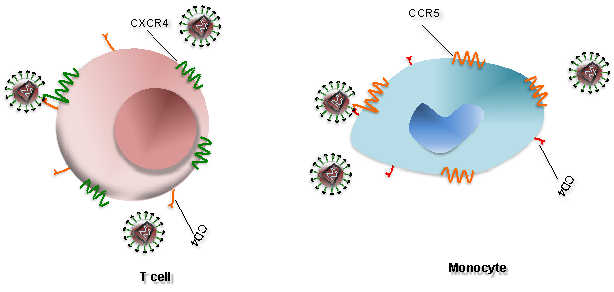
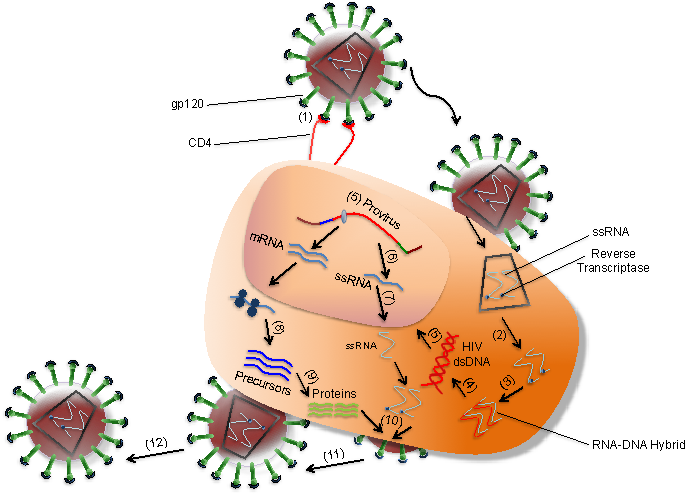
HIV infects T cells via high-affinity interaction between the virion envelope glycoprotein (gp120) and the CD4 molecule. The infection of T cells is assisted by the T-cell co-receptor called CXCR4 while HIV infects monocytes by interacting with CCR5 co-receptor (Figure 1). As illustrated in Figure 2, after gp120 binds to CD4 on the T cell (1). Nucleocapsids containing viral genome and enzymes enters the target cell (2). Following the release of viral genome and enzymes from the core protein, viral reverse transcriptase catalyses reverse transcription of ssRNA to form RNA-DNA hybrids (3). To yield HIV dsDNA the viral RNA template is partially degraded by ribonuclease H and the second DNA strand is synthesized (4). The viral dsDNA is translocated into the nucleus and integrated into the host genome by the viral integrase enzyme (5). Transcription factors transcribe the proviral DNA into genomic ssRNA (6), which is exported to cytoplasm (7). In the cytoplasm, host-cell ribosomes catalyse synthesis of viral precursor proteins (8). The viral precursor proteins are cleaved into viral proteins by viral proteases (9). HIV ssRNA and proteins assemble beneath the host-cell plasma membrane (10) forming virion buds from it (11). Maturation occurs either in the forming buds or after budding from the host cell (12). During maturation, HIV proteases cleave the poly-proteins into individual functional HIV proteins. The mature virions are able to infect another host cell.
Innate immune response to HIV
Innate immune cells (e.g., dendritic cells and natural killer cells) are the first line of defence which HIV encounters upon entry to the body.
Macrophages. Tissue macrophages are one of the target cells for HIV. These macrophages harbour the virus and are known to be the source of viral proteins. However, the infected macrophages are shown to lose their ability to ingest and kill foreign microbes and present antigen to T cells. This could have a major contribution in overall immune dysfunction caused by HIV infection.
Dendritic cells (DCs). DCs are large cells with dendritic cytoplasmic extensions. These cells present processed antigens to T lymphocytes in lymph nodes. Epidermal DCs, expressing CD1a and Birbeck granules, are probably among the first immune cells to combat HIV at the mucosal surfaces. These cells transport HIV from the site of infection to lymphoid tissue. The follicular DCs, found in lymphoid tissue, are also key antigen-presenting cells that trap and present antigens on their cell surfaces. In the lymph node follicles, DCs provide signals for the activation of B lymphocytes.
Natural killer (NK) cells. NK cells have lytic activity against cells that have diminished expression of major histocompatibility complex (MHC) I antigens. Because the presence of MHC class I is required for peptide presentation to T cell receptors, NK cells are important line of defence when HIV escapes the cellular immune response. NK cells proliferate in response to type 1 interferon secreted by DCs. These stimulated NK cells release cytokines such as interferon γ (IFN-γ), tumour necrosis factor α (TNF-α), and chemokines to activate T-cell proliferation (cellular immune response). NK cells also inhibit viral replication by releasing IFN-γ.
Adaptive immune response to HIV
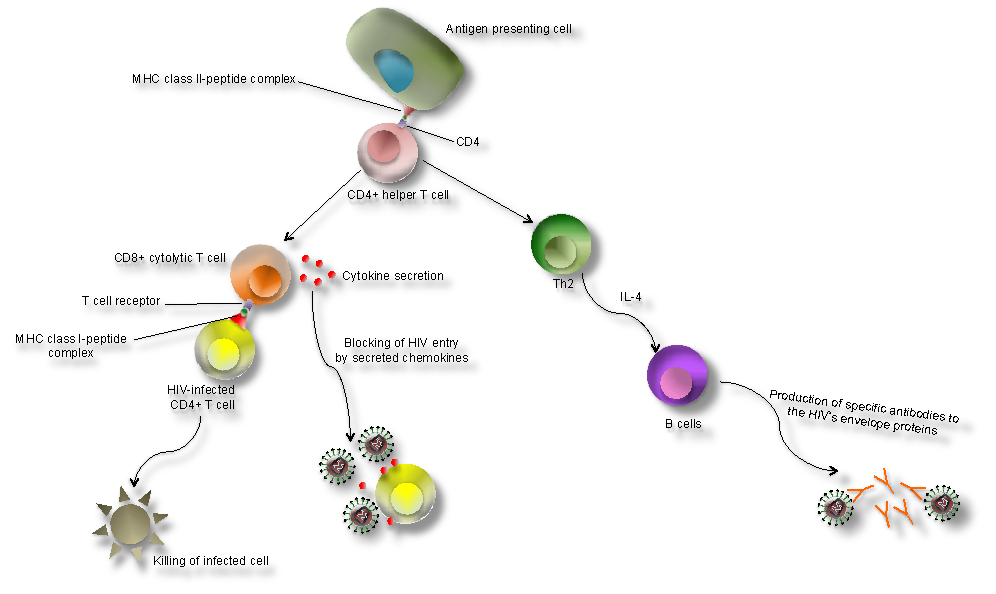
Cellular immune response to HIV. The cellular immune response is induced upon the entry of HIV into the target cells (e.g., T cells) and synthesis of viral proteins (Figure 1). MHC class I on the cell surface displays the intracellularly degraded HIV peptide fragments for recognition by T-cell receptors (TCR) on CD8+ T cells (Figure 3). CD8+ T cells lyse HIV infected cells and secrete cytokines, i.e. interferon-γ (IFN-γ), tumor necrosis factor α (TNF-α), and chemokines, i.e. MIP-1 α, MIP β and RANTES, that inhibit virus replication and block viral entry into CD4+ T cells. Development of CD8+ T cells is crucial for control of HIV replication. This results in declining viraemia after primary infection. In the early stages of infection, CD4+ T cells lose their proliferative capacity and therefore their contribution to viral control is minor. However, during chronic infection CD4+T cells are present and secrete interleukin-2 (IL-2) or cytokines, such as IFN-γ, to control viraemia.
Humoral response to HIV. The humoral immune response occurs later in infection; therefore, the level of antibodies during the acute infection is very low. Non-neutralising antibodies to structural proteins (i.e. P17 and P24) are first to appear and generally do not persist. Later neutralising antibodies specific to proteins, involved in the entry of the virus into the cells, will be generated. These antibodies are specific to: (1) the variable region of gp120 (V3); (2) CD4 binding sites and chemokine receptors (i.e., CXCR4 and CCR5); (3) the transmembrane protein gp41. Potent neutralizing antibodies have been shown to play a major role in controlling HIV infection in a few symptom-free HIV+ individuals who maintain high level of CD4+ T cells and low viral load.
Why does the immune system fail to fight the HIV virus?
There are various reasons which can contribute to the failure of the immune system to control HIV infection and prevent AIDS development. By infecting CD4+ T cells, HIV is able to replicate predominantly in activated T cells and paralyse one of the main components of adaptive immune system. HIV can also establish latent infection in CD4+ T cells and remain invisible to CD8+ T cells and therefore replication can occur later in the infection and generate new virions. Antigenic mutation within the T-cell epitopes can affect the binding capacity of MHC molecules to the viral peptides, resulting in the inability of the TCRs to recognise the MHC-peptide complex. Finally, HIV is able to hide from anti-HIV antibodies by expressing non-immunogenic glycans on key antibody epitopes.
© The copyright for this work resides with the BSI.