BiteSized Immunology: Pathogens & Disease

Tuberculosis
Tuberculosis (TB) in humans is caused by infection with Mycobacterium tuberculosis, which is a non-motile, slow-growing, rod-shaped bacillus. Current figures from the World Health Organisation estimate 8.8 million people developed active TB disease globally in 2010, leading to approximately 1.45 million deaths as a result of the infection.
Tuberculosis is spread via respiratory droplets that contain the tubercle bacillus. These are expelled from individuals with active TB and subsequently inhaled by contacts. While most droplets will be stopped from entering the body by the physical barriers found in the upper respiratory tract, those that are less than 1-2 µm in size are able to bypass these barriers and reach the lower respiratory tract and lungs. It is here that the bacteria encounter cells of the immune system and the battle between host and pathogen begins.
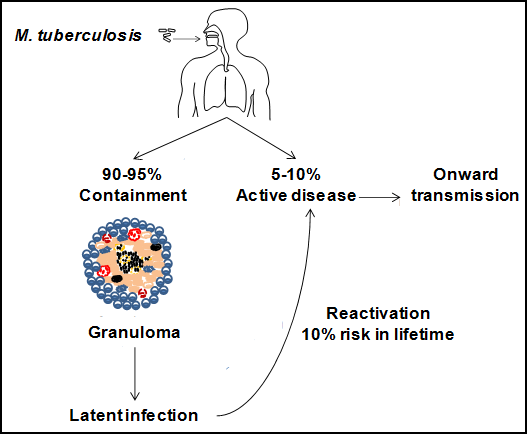
The outcome of infection is dependent on the protective power of the host’s immune system and the pathogenicity of the bacteria. The majority of individuals will be able to control infection and contain it within a granuloma, which is an aggregate of immune cells that walls off the mycobacteria, but does not eradicate them. As bacteria are still present, the infection is described as latent and the individual is at risk of future reactivation of disease if they become immunocompromised.
The first cells encountered by the mycobacteria in the lung are the alveolar macrophages. These are phagocytes, which respond to invading pathogens in a non-specific manner and provide an initial line of defence. Macrophages and other antigen presenting cells (APCs) recognise and phagocytose the mycobacteria, engulfing them into a phagosome.
Typically upon phagocytosis, the bacteria-containing phagosome matures and fuses with endosomes and lysosomes. The environment becomes acidic and nutrient-poor, and the bacteria are exposed to antimicrobial peptides and degrading lysosomal enzymes, such as lysozyme. Pathogenic mycobacteria have developed mechanisms to subvert the host defences by blocking phagosomal maturation in resting macrophages. However, activation of macrophages by the cytokine interferon-γ (IFNγ), promotes bacterial killing via the formation of toxic reactive oxygen intermediates (ROI) and reactive nitrogen intermediates (RNI). Activated macrophages also release an array of cytokines and chemokines, including tumour necrosis factor α (TNFα), which induce a pro-inflammatory response and direct immune cells to the site of infection.
APCs known as dendritic cells, together with activated macrophages, are able to process antigen and present components on their surface in conjunction with MHC class II molecules. Dendritic cells migrate to draining lymph nodes where they encounter large numbers of naive T cells. Naive CD4+ T cells sample antigen/MHC complexes on the surface of migrating APCs. After recognising the antigen/MHC complex specific for its T-cell receptor (TCR), the CD4+ cells become activated, proliferate and, in the presence of proinflammatory cytokines such as IFNγ and IL12, differentiate into T helper (Th)-1 cells.
The Th1 effector cells migrate back to the lungs via chemokine gradients produced by inflammation at the site of infection, where these activated T cells interact with MHC/antigen complexes on the surface of infected macrophages and produce a range of cytokines including IFNγ; leading to further activation of macrophages and triggering the potent antimicrobial activities of the primed Th1 cells.
An orchestrated combination of innate and Th1-dominant adaptive immune responses culminates in the development of granulomas. The infiltration of cells into the lungs during the early innate response becomes organised into a primary granuloma with centrally located macrophages. This leads to the formation of a larger well-organised solid granuloma when adaptive immunity is initiated with the infiltration of specific T lymphocytes and also CD8+, NK and γδT cells. Macrophages at the centre will often be infected, have an activated appearance or be differentiated into epitheloid cells. Some also combine to form giant multinucleated cells.
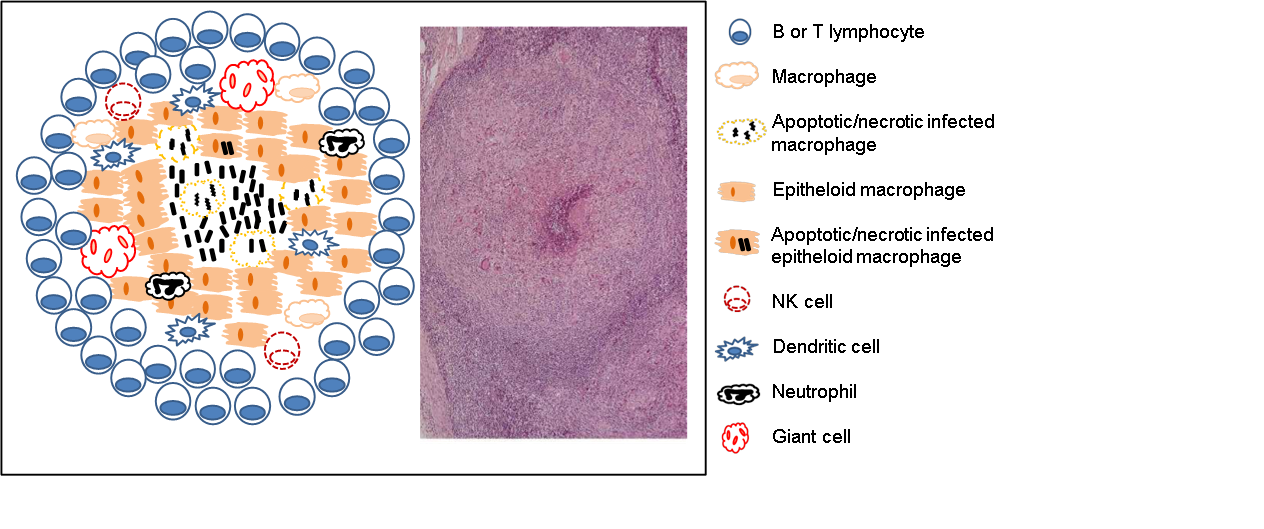
The centre of the granuloma may exhibit caseous necrosis and have a cheese-like appearance. If the infection continues, the centre may liquefy, producing an environment in which the bacteria can grow extracellularly. Cavitation may occur if the liquefied contents are released into the bronchial tree where they can then be expelled externally and the infection transmitted to others. More commonly, the granulomas will undergo fibrosis or calcification and the infection is contained and becomes latent.